Развитию центральной нервной системы ребенка способствуют гормоны

Центральной нервной системе (ЦНС) принадлежит ведущая роль в организации адаптационных процессов, протекающих в ходе индивидуального развития. Поэтому динамика морфо-функциональных преобразований в этой системе скачивается на характере деятельности всех систем организма.
Количество нейронов ЦНС достигает максимального количества у 24-недельного плода и остается постоянным до пожилого возраста. Дифференцированные нейроны уже не способны к делению, и постоянство их численности играет основную роль в накоплении и хранении информации. Глиальные клетки продолжают оставаться незрелыми и после рождения, что обусловливает дефицит их защитной и опорной функций для ткани мозга, замедленные обменные процессы в мозге, его низкую электрическую активность и высокую проницаемость гемато-энцефалического барьера.
К моменту рождения мозг плода характеризуется низкой чувствительностью к гипоксии, низким уровнем обменных процессов (метаболизма) и преобладанием в этот период анаэробного механизма получения энергии. В связи с медленным синтезом тормозных медиаторов в ЦНС плода и новорожденного легко возникает генерализованное возбуждение даже при небольшой силе раздражения. По мере созревания мозга активность тормозных процессов нарастает. На ранних стадиях внутриутробного развития нервный контроль функций осуществляется преимущественно спинным мозгом. В начале плодного периода (восьмая—десятая недели развития) появляется контроль продолговатого мозга над спинным. С 13—14 недели появляются признаки мезенцефального контроля нижележащих отделов ЦНС. Корригирующие влияния коры на другие структуры ЦНС, механизмы, необходимые для выживания после рождения, выявляются в конце плодного периода. К этому времени определяются основные типы безусловных рефлексов: ориентировочный, защитный (избегание), хватательный и пищевой. Последний, в виде сосательных и глотательных движений, наиболее выражен.
Развитию ЦНС ребенка в значительной мере способствуют гормоны щитовидной железы. Снижение выработки тиреоидных гормонов в фатальном или раннем постнатальном периодах приводит к кретинизму в связи с уменьшением числа и размеров нейронов и их отростков, нарушением метаболизма в мозге белка и нуклеиновых кислот, а также передачи возбуждения в синапсах.
В сравнении со взрослыми дети имеют более высокую возбудимость нервных клеток, меньшую специализацию нервных центров. В раннем детстве многие нервные волокна еще не имеют миелиновой оболочки, обеспечивающей изолированное проведение нервных импульсов. Вследствие этого процесс возбуждения легко переходит с одного волокна на другие, соседние. Миелинизация большинства нервных волокон у большинства детей заканчивается к трехлетнему возрасту, но у некоторых продолжается до 5—7 лет. С плохой «изоляцией» нервных волокон во многом связана высокая иррадиация нервных процессов, а это влечет за собой несовершенство координации рефлекторных реакций, обилие ненужных движений и неэкономичное вегетативное обеспечение. Процессы миелинизации нормально протекают под влиянием тиреоидных и стероидных гормонов. По мере развития, «созревания» нейронов и межнейронных связей, координация нервных процессов улучшается и достигает совершенства к 18—20 годам.
Возрастные изменения функций ЦНС обусловлены и другими морфологическими особенностями развития. Несмотря на то, что спинной мозг новорожденного является наиболее зрелой частью ЦНС, его окончательное развитие завершается одновременно с прекращением роста. За это время его масса увеличивается в 8 раз.
Основные части головного мозга выделяются уже к третьему месяцу эмбрионального периода, а к пятому месяцу эмбриогенеза успевают сформироваться основные борозды больших полушарий. Наиболее интенсивно головной мозг человека развивается в первые 2 года после рождения. Затем темпы его развития немного снижаются, но продолжают оставаться высокими до 6—7 лет, когда масса мозга ребенка достигает 80% массы мозга взрослого.
Головной мозг развивается гетерохронно. Быстрее всего идет созревание стволовых, подкорковых и корковых структур, регулирующих вегетативные функции организма. Эти отделы по своему развитию уже в 2—4 года похожи на мозг взрослого человека [4]. Окончательное формирование стволовой части и промежуточного мозга завершается только в 13—16 лет. Парная деятельность полушарий головного мозга в онтогенезе меняется от неустойчивой симметрии к неустойчивой асимметрии и, наконец, к устойчивой функциональной асимметрии. Клеточное строение, форма и размещение борозд и извилин проекционных зон коры приобретают сходство со взрослым мозгом к 7 годам. В лобных отделах это достигается только к 12 годам. Созревание больших полушарий полностью завершается только к 20—22 годам.
В возрасте 40 лет начинаются процессы дегенерации в ЦНС. Возможна демиелинизация в задних корешках и проводящих путях спинного мозга. С возрастом падает скорость распространения возбуждения по нервам, замедляется синаптическое проведение, снижается лабильность нервных клеток. Ослабляются тормозные процессы на разных уровнях нервной системы. Неравномерные, разнонаправленные изменения в отдельных ядрах гипоталамуса приводят к нарушению координации его функций, изменениям в характере вегетативных рефлексов и в связи с этим к снижению надежности гомеостатического регулирования. У пожилых людей снижается реактивность нервной системы, ограничиваются возможности адаптации организма к нагрузкам, хотя у отдельных лиц и в 80 лет функциональное состояние ЦНС и уровень адаптационных процессов могут сохраняться такими же, как и в среднем зрелом возрасте. На фоне общих изменений в вегетативной нервной системе наиболее заметно ослабление парасимпатических влияний.
Центральная нервная система является наиболее устойчивой, интенсивно функционирующей и долгоживущей системой организма. Ее функциональная активность обеспечивается длительным сохранением в нервных клетках нуклеиновых кислот, оптимальным кровотоком в сосудах мозга и достаточной оксигенацией крови. Однако при нарушении этих условий функциональные возможности ЦНС резко уменьшаются.
Источник
У
плода человека количество нейронов ЦНС
достигает максимума в
первые 20-24 недели внутриутробного
развития и остается постоянным
до пожилого возраста. После рождения
дифференцированные нейроны
уже не обладают митотической активностью,
тогда как пролиферация
глиальных клеток мозга продолжается.
Постоянство
183



 числа
числа
нейронов головного мозга играет основную
роль в накоплении
и сохранении информации.
Незрелость
глиальных клеток обуславливает дефицит
защитной и
опорной
функций для ткани мозга, замедленные
обменные процессы в
мозге и его низкую электрическую
активность, медленное и неэкономичное
распространение возбуждения по нервным
волокнам (из-за
недостаточной миелинизации), а также
большую проницаемость
гематоэнцефалического барьера (ГЭБ).
Во
время рождения ребенка его мозг
характеризуется низкой чувствительностью
к гипоксии, низким метаболизмом и
преобладанием в этот период анаэробного
пути получения энергии. Относительные
величины массы мозга новорожденного и
размеров его головы
в 4 раза больше, чем у взрослого человека.
У плода и новорожденного
в связи с низким уровнем тормозных
медиаторов в ЦНС
легко возникает генерализованное
возбуждение даже при очень небольшой
силе раздражения. Активность тормозных
процессов по мере
созревания мозга нарастает.
Процессы
возбуждения и торможения можно выявить
на ЭЭГ в последней
фазе внутриутробного периода, причем
раньше в спинном мозге,
а затем в коре больших полушарий,
вызванные потенциалы в
незрелом мозге отличаются от аналогичных
в зрелом мозге: низкая
амплитуда, удлиненный латентный период,
нестабильность формы
и даже исчезновение ответа при повторных
стимулах. Эти критерии
имеют важное практическое значение для
определения степени
функциональной зрелости мозга ребенка.
Электрическая
активность мозга. Спонтанная
электрическая активность
мозга появляется в раннем пренатальном
периоде. ЭЭГ может быть
отведена от мозга плода путем наложения
электродов на живот матери
или при введении их во влагалище, а в
случае преждевременных родов
(например, после кесарева сечения) —
прямо с поверхности
головы. Биотоки начинают регистрироваться
у плода с 3-5 месяцев,
хотя кора головного мозга появляется
лишь с 6-го месяца. Следовательно,
биотоки мозга, проявившиеся во время
раннего фе-тального
периода, связаны с субкортикальными
структурами.
Тип
ЭЭГ, свойственный здоровым новорожденным
в период бодрствования,
формируется к 36-й неделе внутриутробного
развития. Начиная
с этого периода, ЭЭГ приобретает
непрерывный характер, она
синергична в обоих полушариях;
регистрируются волны низкой амплитуды
с частотой 1-2 и 3-4 Гц. Типичная картина
сна появляется
на ЭЭГ в 7 или 8 месяцев пренатальной
жизни. Преобладающей
формой сна недоношенных детей является
активный сон, во
время которого отмечаются десинхронизация,
быстрые движения глаз,
вздрагивание, отсутствие шейных
тонических рефлексов, нерегулярный
сердечный и дыхательный ритм. По мере
развития плода продолжительность
быстрого сна снижается, а доля медленного
сна увеличивается.
Динамика ЭЭГ показана на рис. 18.8.
Свойственный человеку
а-ритм ЭЭГ отмечается с 3-4 месяца
постнатального развития
и обычно стабилизируется к 13 годам.
184

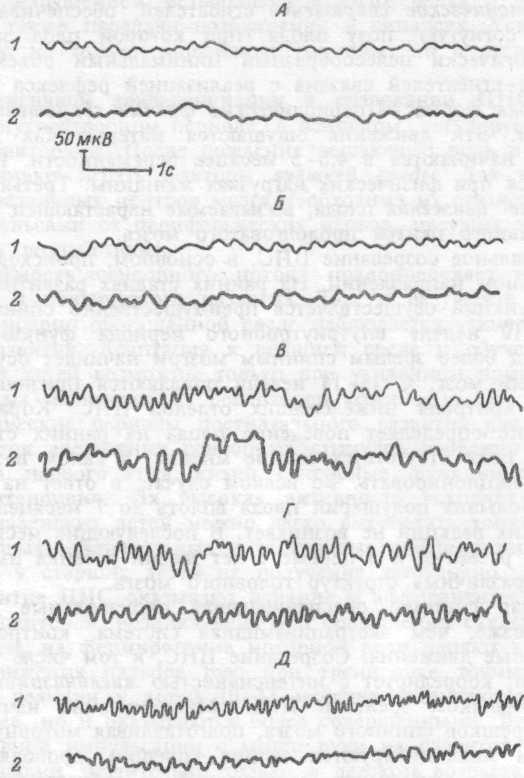
Рис. 18.8. Возрастная
динамика электроэнцефалограммы.
1
— в затылочной, 2 — в центральной
областях. Электроэнцефалограммы:
А — новорожденного, Б
— 3-х месячного ребенка, В — 2-х
летнего ребенка, Г
— 3-х летнего ребенка, Д — 6-ти
летнего ребенка.
Движения
плода и созревание рефлекторных дуг.Так
называемые
спонтанные движения плода появляются
в течение первых трех
месяцев жизни плода. К концу внутриутробного
периода определяются
три основных типа рефлексов: защитный
(избегание),
пищевой
и
хватательный.
Ранние
движения плода могут быть результатом
нескольких причин, но чаще — его
асфиксией, вследствие сдавления
маткой пупочного канатика (возможно,
движения плода служат
восстановлению газообмена).
185

 Для
Для
плода свойственны три формы активности
скелетных мышц. Первая
— тоническое
сокращение сгибателей,
обеспечивающее характерную
«согнутую» позу плода, при которой
плод занимает в матке
биологически целесообразный минимальный
объем. Активность мышц-сгибателей
связана с реализацией рефлекса на
растяжение.
Вторая форма — периодические фазные
сокращения мышц-разгибателей.
Эти движения ощущаются матерью как
шевеление плода.
Они начинаются в 4,5-5 месяцев беременности.
Их частота увеличивается
при физических нагрузках женщины. Третья
форма — «дыхательные»
движения плода,
вызываемые нарастающей активностью
дыхательного центра продолговатого
мозга.
Функциональное
созревание ЦНС, в основном, происходит
в кау-до-краниальном
направлении. На ранних стадиях развития
нервный контроль
функций осуществляется преимущественно
спинным мозгом.
К 7-10 неделе внутриутробного периода
функциональный контроль
над более зрелым спинным мозгом начинает
осуществлять продолговатый
мозг. С 13-14 недели появляются признаки
мезен-цефального
контроля нижележащих отделов ЦНС. Кора
больших полушарий
не определяет поведение плода на ранних
стадиях его развития,
и пока отсутствует единое мнение, в
каком возрасте она начинает
функционировать. Во всяком случае, в
ответ на стимуляцию
коры больших полушарий плода вплоть до
7 месяцев его развития
никаких реакций не возникает. В последующие
месяцы внутриутробного
развития и в первые 5 лет жизни ребенка
имеет место созревание
различных структур головного мозга.
Пирамидная
система, обеспечивающая произвольные
движения, созревает
позже, чем экстрапирамидная система,
контролирующая непроизвольные
движения. Созревание ЦНС, в том числе
психических
функций, коррелирует с интенсивностью
миелинизации,
которая
является
признаком зрелости ЦНС. Миелинизация
начинается с передних
корешков спинного мозга, подготавливая
моторную активность,
затем миелинизируются задние корешки,
проводящие пути спинного
мозга, афференты акустической и
лабиринтной систем, необходимые для
постуральной активности позднего
фетального периода.
В головном мозге процесс миелинизации
происходит в первые
2 года жизни ребенка, но продолжается и
позже: у подростка и
даже взрослых людей.
В
течение 3-х последних месяцев внутриутробного
развития у плода
формируются механизмы, необходимые для
выживания новорожденного
ребенка: начинает реализовываться
корковая регуляция, у
такого ребенка уже появляются защитные
и пищевые рефлексы; рефлексы
с мышц и кожи становятся более
локализованными и целенаправленными.
Неонатальный период доношенного ребенка
характеризуется
преимущественной активностью
мышц-сгибателей, а также
— в отличие от недоношенных новорожденных
— относительно
стабильными механизмами терморегуляции.
Развитию
ЦНС ребенка в значительной мере
способствуют гормоны
щитовидной железы. Снижение
выработки тиреоидных гормонов в
фетальном или раннем постнатальном
периодах приводит к кре-
186

 тинизму
тинизму
в связи с уменьшением числа и размеров
нейронов и их отростков,
нарушением метаболизма в мозге белка
и нуклеиновых кислот,
а также передачи возбуждения в синапсах.
Ускоряют ми-елинизацию
не только тиреоидные гормоны, но и
стероидные.
Роль
сенсорной афференциации в созревании
ЦНС. Развитие
ЦНС
во внутриутробном периоде, в основном,
регулируется генетическими
факторами. После рождения решающую роль
в созревании ЦНС
начинают играть факторы внешней
среды. Для
нормального развития
различных центров мозга необходима их
стимуляция нервными
импульсами от периферических нервов,
несущих информацию о
внешних воздействиях.
Интенсивность
сенсорного потока предопределяет и
онтогенез поведения
и психического развития. Так, в результате
воспитания детей
в сенсорно обогащенной среде наблюдается
ускорение психического
развития. Адаптация к внешней среде и
обучение слепоглухонемых
детей возможны только при усиленном
притоке в ЦНС афферентных
импульсов от рецепторов кожи.
В
критические периоды постнатального
развития для созревания мозга
приток адекватных сенсорных стимулов
особенно необходим. У
ребенка первого года жизни сенсорные
функции развиваются весьма интенсивно.
Их высокая активность ускоряет созревание
ЦНС.
У маленьких детей можно легко выработать
тонкие зрительные,
слуховые, обонятельные и тактильные
дифференцировки, в то время
как у старших детей их получение
затруднено.
На
развитие ЦНС оказывают влияние и
афферентные импульсы с проприорецепторов,
возникающие при сокращении скелетных
мышц. В
частности, на формирование моторной
речи влияют координированные
движения пальцев рук: при тренировке
точных движений голосовые
реакции у детей 10-13 месяцев развиваются
не только интенсивнее,
но и оказываются более совершенными.
Влияние про-приоцептивной
импульсации с мышц руки на развитие
коры больших
полушарий значительно только в детском
возрасте, пока идет формирование
речевой моторной зоны мозга.
В
процессе онтогенетического развития
головного мозга имеет место
усиление межполушарного взаимодействия.
У маленьких детей этому
способствует бимануальная координация,
т.е. сочетанные, целенаправленные
движения двумя руками; улучшение
межполушар-ных
связей обусловлено ускорением миелинизации
нервных волокон мозолистого
тела мозга.
В первые 4- 5 лет
жизни происходит быстрое развитие ЦНС:
не только резко прирастает масса мозга,
но отмечается также интенсивная
структурная дифференцировка коры
больших полушарий.
В
раннем постнатальном периоде вначале
образуются вегетативные
условные рефлексы (например, пищевой
рефлекс начинает вырабатываться
со 2-й недели жизни ребенка), затем
появляются двигательные
и, наконец, речедвигательные условные
рефлексы. Скорость
образования натуральных условных
рефлексов постепенно увеличивается.
Искусственные условные рефлексы
появляются позже
187




 натуральных.
натуральных.
Первыми среди них возникают слуховые
и вестибулярные, далее — обонятельные,
вкусовые и т.д.
Основная
онтогенетическая направленность
функционального созревания
мозга ребенка проявляется в возрастающей
специализации корковых
областей и их интеграции, в которой
ведущую роль приобретает
пластичное взаимодействие нервных
центров. Расширение и перестройка
межполушарных и внутриполушарных
связей, в частности,
обеспечивают пятикратное увеличение
площади поверхности мозга
человека от новорожденного до взрослого
человека.
В
процессе онтогенетического развития
мозга имеет место постепенный
переход от первичной локальности реакций
через фазу генерализации
к специализации и интеграции мозговых
структур. Например,
в период новорожденности в ответ на
зрительный стимул на
ЭЭГ в проекционной зрительной области
коры больших полушарий
регистрируется локальный
позитивно-негативный комплекс,
отражающий
приход в кору информации по специальному
каналу. Специфика
зрительного восприятия новорожденных
состоит в его ограниченности
простыми ощущениями. Уже в течение
первых месяцев
жизни у ребенка начинает складываться
активный характер восприятия,
проявляющийся в развитии зрительно-моторной
координации:
фиксация взора, зрительное прослеживание
движущегося объекта,
схватывание его рукой. В основе активного
характера восприятия
лежит взаимодействие проекционного
и непроекционных отделов
коры больших полушарий, что устанавливается
по ЭЭГ. В течение
первых лет жизни ответы на зрительную
стимуляцию широко
распростаняются по коре больших полушарий
в виде сходных вызванных
потенциалов (фаза генерализации). В 6-7
лет проявляются
признаки усиления специализации
различных областей коры в ответ
на поступление в мозг различной зрительной
информации, что приводит
к улучшению распознавания сложных
абстрактных изображений.
Основой
формирования межцентрального
взаимодействия является
развитие
системы горизонтальных внутрикорковых
связей. Важная роль
в обеспечении межцентральной корковой
интеграции принадлежит
а-ритму ЭЭГ. Эмоциогенность воздействия
связана с усилением
процессов межполушарной интеграции. В
силу многофакторности
воздействия мощным и при том весьма
физиологичным методом интеграции
активности мозга являются детские
подвижные игры.
Через игру адекватно усиливается
нагрузка на систему восприятия;
с ее помощью развиваются ассоциативные
связи.
Формированию
интегративной
деятельности ЦНС
новорожденного
предшествует установление в процессе
развития морфофункци-ональных
связей между ее многочисленными
структурами. Проявлением
их созревания служит способность мозга
образовывать реци-прокные
взаимоотношения, доминантный очаг,
функциональную систему
и условный рефлекс.
Первый
условный рефлекс появляется у ребенка
на 2-й неделе после
рождения (условный сосательный рефлекс
на «положение кормления»).
К концу 3-го месяца у него вырабатываются
условные
188
рефлексы
на зрительные раздражения,что делает
поведение ребенка более
адекватным. С 5-го месяца жизни условные
рефлексы можно выработать
уже со всех сенсорных систем, и по мере
онтогенетического
развития они вырабатываются легче. К
концу 1-го года слово
приобретает значение сильного и
самостоятельного сигнала. Доминантные
очаги ребенка характеризуются
неустойчивостью, что служит
причиной его рассеянного внимания.
Последовательно
развивается и центральное
торможение: безусловное
торможение проявляется уже в первые
дни жизни, условное —
к 8-9 дню жизни. Раньше других возникает
угасательное торможение,
к 3-4 месяцу — дифференцировочное, затем
условный тормоз
и, наконец, к 5-му месяцу — запаздывательное
торможение.
Типологические
особенности высшей
нервной деятельности у ребенка
6-11 лет обычно не отмечаются, поскольку
сила, подвижность и уравновешенность
процессов возбуждения и торможения в
этот период
часто меняется. Подростковый период
характеризуется повышением
возбудимости ЦНС и ослаблением процессов
торможения;
ухудшается дифференцирование сигналов,
возрастают латентные периоды
условных реакций, усиливается иррадиация
возбуждения.
В
юношеском возрасте продолжается
структурно-функциональное созревание
коры больших полушарий: усложняется
ансамблевая организация
ее элементов, повышается концентрация
нуклеиновых кислот
в нейронах и усиливается метаболизм
нервных клеток. В восприятии
внешней информации увеличивается роль
лобных областей
коры, наряду с межполушарной интеграцией
завершается и специализация полушарий.
Источник

